Impact Factor
- Issue 14; 2026
- Issue 13; 2026
- Issue 12; 2026
- Issue 11; 2026
- Issue 10; 2026
- Volume 16; 2026
- Advance Articles
- Past Issues
- Cover Images
- Cover Suggestion
- Index & Coverage
- Special Issues
Introduction
Engineering EVs as smart...
Applying EV-based...
Challenges and perspectives in...
Conclusions
Abbreviations
Acknowledgements
References
International Journal of Biological Sciences
International Journal of Medical Sciences
 Global reach, higher impact
Global reach, higher impact

Theranostics 2020; 10(18):8111-8129. doi:10.7150/thno.47865 This issue Cite
Review
Extracellular vesicle-based Nanotherapeutics: Emerging frontiers in anti-inflammatory therapy
Tao-Tao Tang, Bin Wang, Lin-Li Lv, Bi-Cheng Liu ![]()
Institute of Nephrology, Zhong Da Hospital, Southeast University School of Medicine, Nanjing, China.
Received 2020-5-7; Accepted 2020-6-19; Published 2020-7-9
Abstract

Dysregulated inflammation is a complicated pathological process involved in various diseases, and the treatment of inflammation-linked disorders currently represents an enormous global burden. Extracellular vesicles (EVs) are nanosized, lipid membrane-enclosed vesicles secreted by virtually all types of cells, which act as an important intercellular communicative medium. Considering their capacity to transfer bioactive substances, both unmodified and engineered EVs are increasingly being explored as potential therapeutic agents or therapeutic vehicles. Moreover, as the nature's own delivery tool, EVs possess many desirable advantages, such as stability, biocompatibility, low immunogenicity, low toxicity, and biological barrier permeability. The application of EV-based therapy to combat inflammation, though still in an early stage of development, has profound transformative potential. In this review, we highlight the recent progress in EV engineering for inflammation targeting and modulation, summarize their preclinical applications in the treatment of inflammatory disorders, and present our views on the anti-inflammatory applications of EV-based nanotherapeutics.
Keywords: Extracellular vesicles, Anti-inflammatory therapy, Drug delivery, Inflammatory disease, Biotherapy
Introduction
Inflammation underlies a wide range of physiological and pathological conditions [1-4]. In infection or trauma, inflammation is essential to eradicate the noxious pathogens and heal the injured tissues to restore homeostasis. However, if the host fails to eliminate the initiating stimuli or generates a frustrated resolution response, the unresolved inflammation can cause organ dysfunction and become detrimental to the host. In such cases, chronic inflammation ensues and drives the development and progression of various noncommunicable diseases, such as cancer, cardiovascular disease and metabolic disease, which account for 70% of all deaths worldwide [5,6]. Thus, considering inflammation as the cornerstone of many acute and chronic pathological processes, anti-inflammatory therapy is of paramount importance to improve patients' survival rate and survival quality.
With considerable progress in our understanding of the cellular and molecular mechanisms of inflammatory response, therapeutic agents varying from chemical compounds to gene drugs have been developed [7]. However, their clinical translation is often limited by potential side effects, a slow onset of action, or a lack of efficacy, which might be attributed to the poor bioavailability, or to off-target effects after systemic administration [8,9]. Therefore, advanced delivery strategies should be developed to improve pharmacokinetic properties and limit unwanted biological effects in order to realize the full translational potential of these potent drug candidates, particularly for gene and biologic therapies.
There has been growing interest in utilizing biologic nanocarriers by way of cell-derived membranous structures to develop innovative theranostics, and extracellular vesicles (EVs) present exciting potential opportunities and translational possibilities in this area because of distinct advantages such as low immunogenicity, biological barrier permeability, and intrinsic cell targeting properties [10-15]. Based on the current knowledge of their size and biogenesis, EVs can be broadly divided into exosomes, microvesicles, and apoptotic bodies. Here, we focus on the first two classes: exosomes (30-150 nm in diameter) and microvesicles (50-1,000 nm in diameter), which originate from the endosomal system or are shed directly from the plasma membrane, respectively [16,17]. Although EVs were initially regarded as cell dust with no biological significance, their implications in intercellular communication by transferring particular lipid, protein and nucleic acid contents between cells have been well documented recently [18-21]. Importantly, their capacity to transfer cargos has introduced the possibility of using EVs as “Trojan horses” to deliver a variety of agents, including chemotherapeutics, miRNAs, siRNAs, proteins and even nanoparticles [10-12].
Diverging from existing reviews, we will focus on the current state of the art in manufacturing EVs as smart nanotherapeutics for anti-inflammatory therapy. We begin by taking stock of the latest advances in EV engineering technologies, with particular emphasis on their application in inflammation targeting and modulation. Then, we assemble emerging EV-based nanotherapeutics in the treatment of inflammatory diseases. Finally, we discuss the concerns and perspectives regarding the anti-inflammatory applications of EV-based nanotherapeutics.
Engineering EVs as smart nanotherapeutics to target inflammation
Today, a broad spectrum of new drugs is being developed for the relief of inflammation. However, a sizable percentage of them fail to achieve the anticipated effect, in part because conventional therapies are not specific to the diseased tissue or because of the poor pharmacokinetics. EVs are highly tunable and can be engineered and customized for the targeted delivery of different kinds of cargo, which holds promise for reducing off-target effects, decreasing unwanted toxicities, and thereby enhancing a drug's therapeutic efficacy. Thus, employing EVs as a versatile drug carrier may advance and revive multitudinous therapeutic agents with inflammation-combating properties. This section will first consider the recent progress in the techniques of surface engineering and cargo loading, then discuss their applications in developing EV-based anti-inflammatory nanotherapeutics.
Surface engineering of EVs to target inflammation
Surface engineering of EVs is an essential step for therapeutic applications because it endows EVs with additional properties, allowing them to accumulate at target sites, escape immunosurveillance, and achieve prolonged circulation time and enhanced intracellular uptake.
Surface engineering techniques
Cell engineering
Direct genetic engineering of the parental cell directly is a valid method for displaying encodable peptides, antibodies and proteins on the surface of EVs. This is performed either by inserting a targeting epitope into a protein domain, or by fusing a targeting sequence to the EV-enriched membrane proteins [22,23]. Concerning the latter, the lysosomal protein lamp2b is widely used [24-27]. For example, neuron targeting RVG peptide and αv integrin-specific RGD (iRGD) peptide have been anchored on EVs through attachment to lamp2b for tissue-specific delivery [24,26]. Despite these successes, it seems critical that peptides inserted to the N-terminus of lamp2b are probably degraded by endosomal proteases prior to surface expression [27]. Alternatively, the targeting epitope can be fused to a desired protein domain, such as the transmembrane domain of platelet-derived growth factor receptor [28], glycosylphosphatidylinositol anchor peptides [29], or the C1C2-domain of lactadherin [30,31], instead of the whole proteins. However, it should be pointed out that these approaches may cause a functional loss of the host protein, and the recombinant construct may trigger immune activation.
Aside from improving targeting capacity, genetic engineering can be applied to increase the circulation time and cellular uptake of EVs. For example, the expression of vascular stomatitis virus-G on EV membranes enhanced their intracellular uptake by recipient cells [32]. EVs engineered to overexpress CD47 exhibited superior escape from phagocytosis by the mononuclear phagocyte system [33].
Chemical modification
Click chemistry is a type of reaction commonly used for the bioconjugation of molecules to delivery systems via covalent bonds [34]. In general, for EV modification, the alkynes are first grafted to the EV membrane and then reacted with azide-tagged molecules of interest via azide-alkyne cycloaddition. Therefore, this type of modification is less likely than others to dissociate away from the EV. Using this strategy, the azide-modified fluorophores [35], the integrin (ITG) αvβ3-specific cRGD peptide [36], or the neuropilin-1-specific RGE peptide [37] have been successfully conjugated onto the EV surface. Of note, the critical alkyne modification may occur on the EV proteins rather than on the amines on membrane phospholipids. As such, the impacts of this modification on EV protein functions need to be comprehensively evaluated.
The hybridization of EVs with the synthetic liposomes is emerging as an alternative strategy to optimize and augment the properties of natural EVs. A freeze-thaw-induced [38] or PEG-induced [39] fusion of EVs and liposomes has been reported. With this strategy, the EV surface can easily be modified by fusion with liposomes embedding peptides or antibodies. Importantly, the hybrid EV/liposomes showed a better stability and half-life in circulation, as well as lower immunogenicity [38,39]. Additionally, aptamers were displayed on EVs to realize a targeted antitumor delivery modality [40,41]. Nanobody-PEG-lipid conjugates were inserted into the EV membrane to increase the target distribution and prolong the circulation time [42]. Moreover, cationic lipids and pH-dependent fusogenic peptides were employed to increase the cellular EV uptake efficiency and cytosolic release of the cargo [43].
In summary, although surface engineering arms EVs with advanced functionality, research on these issues is still in its infancy. Modifications may alter the membrane protein structure, damage EV function, and exacerbate immunogenicity. Moreover, chemical routes require additional purification steps, which complicate the manufacturing process and increase the chance of EV loss. Thus, exploring more convenient, secure and effective functional moieties is necessary for broadening the therapeutic applications of EVs. Various EV functionalization approaches are shown in Figure 1A.
Surface engineering of EV-based nanotherapeutics. (A) Examples of surface functionalization strategies for EV-based nanotherapeutics. (B) Emerging strategies for developing inflammation targeting EV-based nanotherapeutics.
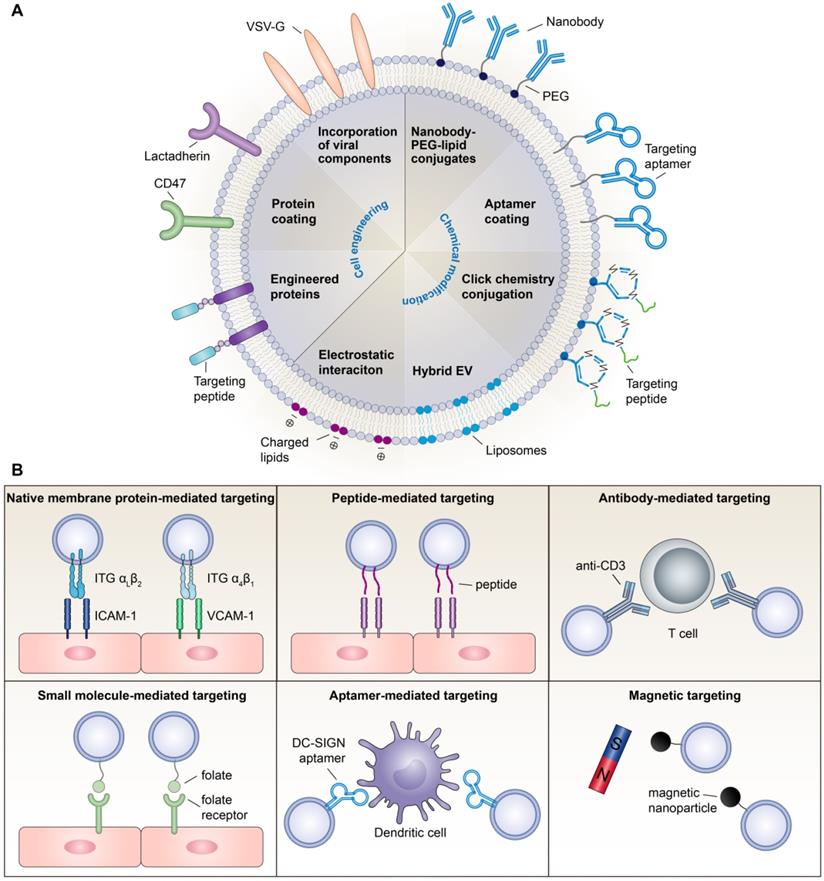
Emerging strategies for creating inflammation-targeting EVs
The homing of EVs towards certain recipient cells is mediated by natural membrane proteins or bioengineered moieties on the EV surface. For anti-inflammatory therapy, targeting EVs to inflammatory cells or inflamed tissues is the primary task.
Natural targeting capacity
There is mounting evidence that the cellular origin of EVs can guide their surface proteins, intrinsic targeting properties, and target cell tropism, so selecting the optimal pair of donor-recipient cells may promote specific EV uptake. For instance, T cell-derived EVs were efficiently taken up by macrophages and microglial cells [44], while EVs from mature dendritic cells were transferred unidirectionally to activated T cells [45]. Recently, macrophage-derived EVs were shown to sense and migrate towards inflamed endothelial cells in both the kidney [46] and the brain [47], which was mediated by the interaction of ITG αLβ2 and ITG α4β1 on EVs with intercellular cell adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1) on endothelial cells, respectively. Similarly, vesicles derived from activated neutrophils can selectively bind inflamed vasculature in the lung and brain because they possess intact targeting molecules of ITG β2 [48-51]. Inspired by the selective recruitment of leukocytes to the site of inflammation [52], leukocyte-derived EVs could facilitate inflammation-targeting drug delivery by the integrin patterns presented on the surface. Nevertheless, the integrin profile of these EVs needs to be further defined. Moreover, immune cell-derived EVs express abundant MHC class I and II molecules [53,54], which may increase the risk of immunogenicity; therefore, a long-term safety assessment is required.
Strategies for active targeting
In addition to native membrane proteins, EVs can be decorated with targeting ligands such as antibodies, peptides, small molecules and aptamers to achieve active targeting. In a recent study, Cheng et al. generated a novel synthetic multivalent antibodies retargeted exosome (SMART-Exo) through cell engineering, which expressed monoclonal antibodies specific for T cell CD3 and cancer cell-associated epidermal growth factor receptor (EGFR) and was shown to induce cross-linking of T cells and EGFR-expressing breast cancer cells [55]. Using this strategy, antibodies for other immune cells or tissue cells are likely to be introduced onto the EV surface, allowing a more specific inflammation targeting. In parallel, targeting peptides have also been used to guide EVs towards lesion sites. For example, Tian et al. conjugated ITG αvβ3-specific cRGD peptides to the exosomal surface by click chemistry, which efficiently targeted to the lesion region of the ischemic brain after intravenous administration [36]. The ischemic myocardium-targeting peptide CSTSMLKAC (IMTP) [56] or cardiac-targeting peptide (CTP) [57] has also been displayed on EVs by fusion with lamp2 protein, conferring the capacity to target the heart. Other inflammation-targeting peptides, such as ICAM-1-binding Cyclo(1,12)PenITDGEATDSGC (cLABL) peptide [58,59] and VCAM-1-binding VHPKQHR peptide [60], have been screened; however, their targeting ability when conjugated to EVs remains to be confirmed.
Alternatively, small molecule-based targeting strategy has also been employed in EV-based therapy. For instance, folate, the most widely used small-molecule targeting agent [61], has been introduced to EVs to target cancer cells that express high levels of folate receptors [41]. Considering that these receptors are also expressed on activated macrophages [62], folate-decorated EVs might be able to target macrophages to function in anti-inflammatory therapy. In addition, aptamers represent another low molecular weight targeting system. A comprehensive review of aptamer-guided EV theranostics in oncology is available elsewhere [63]. With respect to inflammation targeting, a recent study successfully modified EVs with DNA aptamers against an adhesion receptor, ICAM-3 grabbing non-integrin (DC-SIGN), for the targeted delivery of therapeutic RNAs to dendritic cells [64]. However, being nucleic acid-based, degradation in biological media is the most significant challenge of aptamer-targeted systems.
Besides targeting ligand-based strategies, magnetically and optically responsive hybrid-EVs were developed by incubating macrophages with iron oxide nanoparticles and photosensitizers, creating a method that uses magnetic targeting for the tissue-specific delivery of therapeutic EVs [65]. However, magnetic nanoparticles may raise toxicity concerns, and there are difficulties in reaching deep tissues by this method. Overall, strategies for targeting inflammation currently remain limited. More active targeting methods with high affinity to tissue-specific cells need to be explored in the future (Figure 1B).
Cargo engineering of EVs for anti-inflammatory therapy
EVs can be used as carriers for different therapeutic cargos, such as small molecule drugs, proteins, and genetic drugs. Here, the cargo-loading techniques and promising anti-inflammatory agents for EV-based nanotherapeutics will be discussed, with a focus on the improvement of the pharmacokinetics of these drugs delivered by EVs.
Cargo-loading techniques
Exogenous loading
Exogenous loading occurs after EV collection, with the desired therapeutic cargo packaged into EVs by simple coincubation or by chemical or physical approaches, each with its own advantages and limitations (Figure 2). Of note, violent loading procedures that may impact the EV integrity or cause immunogenicity or EV aggregation should be avoided.
Illustration of cargo-loading techniques to produce EV-based nanotherapeutics. Cargo-loading can be carried out either endogenously (pre-loading parental cells with the cargo followed by inducing EV production) or exogenously (directly loading the EVs after their production and isolation).
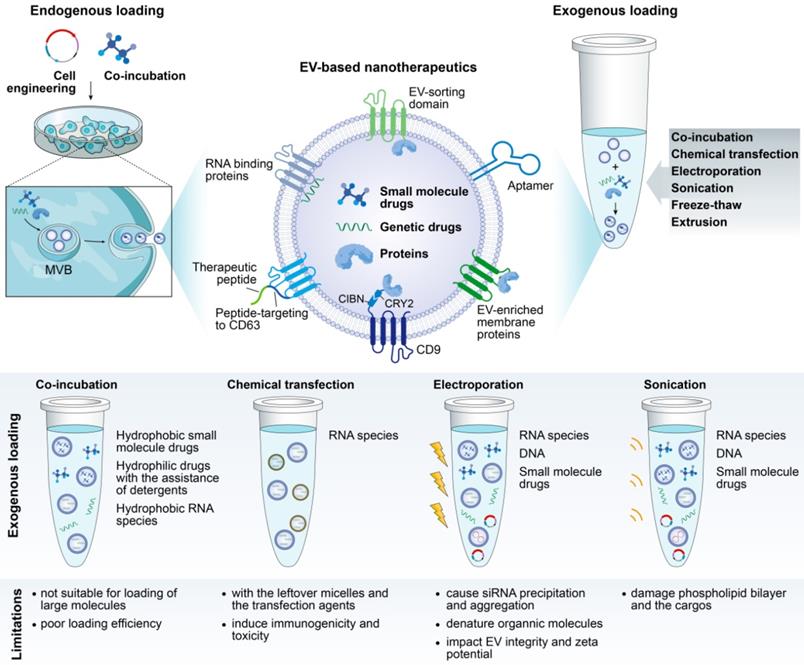
Small hydrophobic molecules can be incorporated into EVs very efficiently by simple incubation [44,66]. However, loading hydrophilic agents in this way requires the assistance of detergents, such as saponin, to loosen the vesicle bilayer [67]. Interestingly, Gao et al. proposed a remote loading of hydrophilic drugs based on the pH gradient between the inside and outside of EVs. Preparing EVs in alkaline buffer to produce a pH gradient tripled the loading efficiency of piceatannol into EVs [49]. Moreover, chemical transfection has been used to load siRNA in EVs by incubating EVs with siRNA-lipofectamine micelles [68]. Despite the loading efficiency, EVs are difficult to purify from the surplus micelles and the transfection agents, which may elicit immunogenetic toxicity.
Physical approaches, such as electroporation, sonication and extrusion, are active loading strategies that generally involve breaking and then restoring EV membrane integrity to enable drug encapsulation. Of these, electroporation has emerged as a favorite method, particularly for RNA species packaging. For example, siRNA/shRNA has been electroporated into EVs to knock down oncogenic mutant KRAS in pancreatic cancer [33]. However, poor loading efficiency has been reported in some cases, which may result from the formation of siRNA aggregates in the process of electroporation [69]. A refinement of siRNA loading with hydrophobically modification was proposed, with convincing and efficient mRNA silencing of human antigen R [70] and huntingtin [71].
Recently, Gao et al. generated an anchor peptide, CP05, that targets CD63 on EVs, enabling direct and effective cargo loading on the surface of EVs. Painting EVs with CP05-conjugated muscle-targeting peptide in combination with CP05-conjugated phosphorodiamidate morpholino oligomer improved dystrophin expression and muscle function in a mouse model of muscular dystrophy [72].
Endogenous loading
Alternatively, the cargo can be endogenously loaded by incubating drugs of interest with parental cells directly or by genetically engineering the parental cells to overexpress a therapeutic RNA or protein, which will then be packaged into the EV cytosol or its membrane (Figure 2). However, how to ensure the selective sorting of desired drugs into EVs remains an open question. At present, a series of RNA binding proteins that specify the loading of miRNA and mRNA into EVs have been identified, including hnRNPA2B1 [73,74], Y-box protein 1 [75], SYNCRIP [76], and ELVA protein HuR [77], which are potential candidates for enabling the controlled loading of therapeutic RNA drugs. Intriguingly, an elegant tool (termed “EXPLORs”) that immobilizes proteins on the inner surface of EVs has been reported [78]. In the EXPLOR producer cells, CRY2 was fused with a cargo protein (CRY2-cargo protein), and CIBN was fused to the N-terminus of an EV marker, CD9 (CIBN-CD9). As a result of the blue light irradiation, CRY2-conjugated cargo proteins were guided to CIBN domains on the surface of early endosomes, thereby effectively introducing cargo proteins into the newly generated EVs [78].
More recently, Yang et al. reported a cellular nanoporation (CNP) method for the scalable production of EVs, along with the loading of therapeutic mRNAs. When cells were transfected with plasmid DNAs by an electrical stimulus on the CNP silicon chip, large quantities of exosomes carrying transcribed mRNAs and targeting peptides were released, producing up to 50-fold more exosomes and an increase of more than 1,000-fold in exosomal mRNA transcripts compared to other strategies [79].
Promising anti-inflammatory agents for EV-based nanotherapeutics
Chemical therapeutics
The loading of small molecular weight drugs into EVs has been extensively studied, including anti-inflammatory drugs such as curcumin [44,66], piceatannol [49], aspirin [80], and dexamethasone [46]. Curcumin is a plant-derived molecule with promising anti-inflammatory activity but suffers from poor solubility. Mixing curcumin with EVs allowed a large amount of drug loading into EVs. Importantly, EV packaging enhanced the solubility, stability and bioavailability of curcumin, which greatly improved its therapeutic efficacy in brain inflammation-related diseases [44,66]. Likewise, dexamethasone is a widely prescribed medication used to treat inflammatory diseases; however, its clinical use is largely restricted due to adverse effects. A recent study prepared dexamethasone in EV formulations, conferring an increased capacity to reduce renal inflammation and fibrosis without significant adverse effects (e.g., hyperglycemia and suppression of HPA axis) during long-term use compared to the free drug [46]. From these studies, EV-mediated delivery may open new opportunities for natural phytochemical compounds or drugs that have poor bioavailability, suboptimal pharmacokinetics, or off-target side effects.
Genetic therapeutics
Throughout the field, delivering nucleic acids, such as mRNA, miRNA and siRNA, within EVs has been widely favored, probably owing to their natural presence as EV cargo. For anti-inflammatory therapy, miRNAs have been successfully loaded into engineered or natural EVs. For example, exogenous miR-155 inhibitors were electroporated into B cell-derived EVs as a gene therapy strategy to relieve LPS-induced inflammation in macrophages. In contrast to conventional transfection methods, EV-mediated miRNA-155 inhibitor delivery resulted in functionally more efficient inhibition of tumor necrosis factor (TNF) production and less cellular toxicity [81]. In addition, it should be noted that various miRNAs are able to modulate inflammation, but whether they are suitable for therapeutic application requires careful assessment. Manipulating miRNA expression may cause unintended biological effects, as the same miRNA can regulate multiple mRNA genes [82].
Unlike miRNA, siRNAs allow the facile and specific inhibition of any gene, and EV-mediated siRNA delivery is demonstrated to be an effective method for RNA interference (RNAi) therapy. To our knowledge, applying EV-siRNA in the fight against inflammation has not yet been reported, but many lessons can be learned from other fields, especially in the area of tumor RNAi therapy [83,84]. First, using the transfection of leukemia cells as an example, EVs achieved remarkable delivery efficiency of antisense oligonucleotides without any toxicity to cells compared to commercial transfection reagents [85]. Additionally, in T cells and monocytes, the gene mitogen-activated protein kinase 1 was selectively silenced by using siRNA-loaded EVs [86]. Since most immune cells are difficult to transfect, the use of EVs as a vector can address this problem with enhanced transfection efficiency but low toxicity. Second, by the right of the modifiability of the EV membrane, EV-based RNAi drugs are more likely to reach the specific diseased tissues. For example, siRNA against BACE1 was encapsulated into EVs modified with neuron targeting RVG peptide, thereby leading to specific gene knockdown in the brain with minimal toxicity and immune stimulation, even during repeated systemic administration [24]. Moreover, the surface expression of endogenous signaling ligands such as CD47 on EVs can increase the systemic circulation half-life and improve cellular uptake [33,87]. Translating these advances into anti-inflammatory therapy may enable new breakthrough treatments. However, EV-based RNAi therapy still faces considerable challenges, including nonspecific gene silencing, limited silencing effects, and potential in vivo genotoxicity.
mRNA-based therapeutics hold great potential for the treatment of inflammation, as they can be exploited for precise and individualized therapy. A recent study demonstrated a proof of concept: implanting engineered producer cells termed EXOtic devices in living mice to generate therapeutic EVs loaded with biopharmaceutical-encoding mRNAs in-situ, which was intended to enhance EV production, specific mRNA packaging, and delivery of the mRNA into the target cells [88]. In models of Parkinson's disease, EXOtic devices successfully ameliorated neurotoxicity and neuroinflammation by delivering catalase mRNA via EVs from implanted cells. Through this technology, we can load a specific mRNA of interest into EVs, and no need of concentration and purification of EVs as the traditional method does may further protect the stability of therapeutic mRNA inside the EVs.
Protein therapeutics
Proteins, such as enzymes, peptides, and cytokines, have become a class of important biotherapeutics for treating diseases because of their high biological activity and superb specificity [89]. EVs have inherent advantages in delivering protein drugs with anti-inflammatory potency. For example, Kou et al. found that mesenchymal stem cells (MSCs) produce and secrete interleukin-1 receptor antagonist (IL-1RA), a natural inhibitor of the proinflammatory cytokine IL-1β, associated with EVs via the Fas/Fap-1/Cav-1 cascade triggered by TNF. IL-1RA-EV contributed to wound healing in both the gingiva and the skin [90]. Another study engineered parental cells to release EVs overexpressing the anti-inflammatory cytokine IL-4 and containing the endogenous “eat me” signal lactadherin (Mfg-e8) on the surface to target phagocytes. A single injection of such EVs into the cisterna magna significantly ameliorated neuroinflammation by inducing the M2 phenotype in the recipient microglia [91]. Recently, IκBα, a super-repressor of nuclear NF-κB activation, was loaded into EVs through the EXPLOR system to attenuate mortality and systemic inflammation in septic mouse models [92]. These findings offer important inspirations that commandeering nature's own anti-inflammatory mechanisms to inhibit inflammation may accelerate the development of EV-based nanotherapeutics. Future therapies may be able to take advantage of other suppressors of inflammation, such as TGF-β and IL-10 family cytokines [93]. Moreover, a kind of decoy EV was generated by presenting the TNF binding domain of human TNF receptor-1 on the EV surface, which antagonized TNF-induced signaling in cellular models of inflammation [94]. In this way, decoy EVs displaying multiple receptors of inflammatory cytokines can be further exploited as biological sponges to absorb detrimental factors in blood or tissues.
Overall, EV-mediated drug delivery presents excellent application prospects in the anti-inflammatory therapy and gives new scope to a very large number of drugs by enhancing their solubility, bioavailability, stability, activity, or safety.
Applying EV-based nanotherapeutics in the fight against inflammatory diseases
Two encompassing therapeutic applications of EVs can be summarized from current research. By virtue of their bioactive components, EVs have intrinsic therapeutic potential in tissue repair and regenerative medicine as well as in immunomodulation. On the other hand, natural or engineered EVs are being utilized as delivery vectors for different cargos of choice. Below, EV-based nanotherapeutics with anti-inflammatory properties and the underlying therapeutic mechanisms will be illustrated with contemporary examples taken from therapy for inflammatory diseases.
EVs as therapeutic biomolecules with intrinsic anti-inflammatory activity
EVs from various sources have therapeutic potency, among which MSC-derived EVs appear particularly useful in the treatment of diverse conditions, including the treatment of inflammatory disorders of the respiratory system [95-98], heart [99-101], liver [102-104], kidney [105-108], nervous system [109-113], arthrosis [114-116], muscle [117-119] and others [120-124] (Table 1). The therapeutic action of MSC-EVs is reliant on their transfer of genetic materials and proteins. However, because most studies of MSC-EVs have focused on their efficacy rather than on their cargos, it is far from clear which anti-inflammatory entity is responsible for any given effect. A recent study delineated the components of IFNγ-activated MSC-derived EVs through deep RNA sequencing and proteomics, revealing that such EVs were rich in anti-inflammatory and neuroprotective RNAs and proteins [111]. This study enhances the understanding of the potential mechanisms by which MSC-EVs exert therapeutic function in multiple sclerosis.
Representative MSC-derived EVs with anti-inflammatory properties
| Indication | EV source and Isolation method | Therapeutic schedule | Effective molecules | Treatment outcome |
|---|---|---|---|---|
| Respiratory | ||||
| Influenza [95] | Swine BM-MSC; Differential centrifugation with ultracentrifugation | 80 μg/kg body weight, single dose; intratracheal | RNAs | -Reduced virus shedding and influenza virus replication -Reduced apoptosis and proinflammatory cytokines -Ameliorated influenza virus-induced acute lung injury |
| Neonatal hyperoxic lung injury [96] | Human UC-MSC; Differential centrifugation with ultracentrifugation | 20 μg, single dose; intratracheal | VEGF protein and mRNA | -Suppressed macrophage activation and proinflammatory cytokines secretion -Improved alveolarization and angiogenesis -Attenuated hyperoxic lung injuries |
| Pneumonia [97] | Human BM-MSC; Differential centrifugation with ultracentrifugation | dosed by total cell count, single dose; intratracheal or intravenous | KGF mRNA | -Reduced inflammatory cell infiltration and cytokines secretion -Increased bacterial clearance -Prolonged survival |
| Allergic airway inflammation [98] | Human iPSC-MSCs; Anion-exchange chromatography | 2×1010 particles, single dose; intravenous | miR-146a-5p | -Inhibited the function of human ILC2s in vitro -Alleviated ILC2 levels, inflammatory cell infiltration and mucus production in the lung, and improved airway hyperresponsiveness |
| Cardiovascular | ||||
| Myocardial I/R injury [99] | Mouse BM-MSC; Differential centrifugation with ultracentrifugation | 20 μg, single dose; myocardial injection | miR-182 | -Converted pro-inflammatory macrophages to M2-like phenotype -Reduced infarct size and alleviated inflammation |
| Myocardial infarction [100] | BM-MSC; ExoQuick-TC (System Bioscience) | 80 μg, single dose; myocardial injection | Not studied | -Promoted angiogenesis and inhibited proliferation of lymphocytes in vitro -Reduced infarct size, and preserved cardiac systolic and diastolic performance |
| Sepsis-induced cardiac injury [101] | Mouse BM-MSC; Differential centrifugation with ultracentrifugation | 2 μg/g body weight, single dose; intravenous | miR-223 | -Reduced polymicrobial sepsis triggered cardiac dysfunction, apoptosis and inflammatory response |
| Hepatic | ||||
| Hepatic I/R injury [102] | Human UC-MSC; Differential centrifugation with ultracentrifugation | 10 mg/kg body weight, single dose; intravenous | MnSOD | -Alleviated neutrophil infiltration and oxidative stress -Protected against hepatic apoptosis and restored liver function |
| Chronic liver failure [103] | Human ESC-MSCs; Differential centrifugation with ultracentrifugation | 350 μg, single dose; intraperitoneal | Not studied | -Hydrogel-mediated delivery improved the anti-fibrosis, anti-inflammation, anti-apoptosis, and regenerative effects of MSC-EVs |
| Acute liver failure [104] | Human and mouse BM-MSC; Differential centrifugation with ultracentrifugation | 2×108~2×1010 particles, single dose; intravenous/ intraperitoneal | Y-RNA-1 | -Reduced hepatic injury, modulated cytokine expression, and increased survival by systemic administration |
| Renal | ||||
| Renal I/R injury [105] | Mouse BM-MSC; Differential centrifugation with ultracentrifugation | 80 μg, single dose; renal capsule injection | Not studied | -Hydrogel-mediated delivery enhanced the anti-apoptosis and anti-inflammatory effects of MSC-EVs -Promoted endothelial cell proliferation and angiogenesis, and inhibited chronic renal fibrosis |
| Metabolic syndrome and renal artery stenosis [106] | Swine AD-MSC; Differential centrifugation with ultracentrifugation | 1×1010 particles, single dose; intrarenal injection | IL-10 | -Induced monocytes to differentiate into M2-like macrophages, and reduced renal inflammation -Ameliorated renal hypoxia and scarring |
| Renal I/R injury [107] | Human UC-MSC; Differential centrifugation with ultracentrifugation | 100 μg, single dose; intravenous | miR-15a miR-15b miR-16 | -Suppressed CX3CL1 expression, macrophage infiltration and cell apoptosis -Improved renal function and renal fibrosis |
| Renal I/R injury [108] | Human UC-MSC; Differential centrifugation with ultracentrifugation | 100 μg, single dose; intravenous | miRNAs | -Improved tubular injury and protected renal functions by modulating NK cells |
| Neurological | ||||
| Cortical injury [109] | Monkey BM-MSC; Differential centrifugation with ultracentrifugation | 4×1011 particles, two doses (Day 1 and 14 post-injury); intravenous | Not studied | -Reduced neuroinflammation and shifted microglia towards an anti-inflammatory phenotype -Recovered motor function |
| Hippocampal damage [110] | Human BM-MSC; Differential centrifugation with ultracentrifugation | 15 μg, two doses; intracardiac injection | Cytokines and factors | -Suppressed extensive inflammation, reactive astrogliosis, and increased integrity of the BBB -Rescued memory and learning deficiencies |
| Multiple sclerosis [111] | Human BM-MSC; Differential centrifugation with ultracentrifugation | 150 μg, single dose; intravenous | RNAs and proteins | -Decreased neuroinflammation and upregulated Tregs -Reduced demyelination and the clinical score of EAE mice |
| Acute spinal cord injury [112] | Human BM-MSC; Tangential flow filtration | 1×109 particles, single dose; intravenous | Not studied | -Diminished inflammatory response with apparent astrocyte and microglia disorganization, and improved functional recovery |
| Preterm brain injury [113] | Human BM-MSC; Differential centrifugation with ultracentrifugation | dosed by total cell count, two doses (3 h before and 24 h after injury); intraperitoneal | Not studied | -Prevented reactive astrogliosis and microgliosis -Reduced neuronal cell death, and restored white matter microstructure |
| Musculoskeletal | ||||
| Osteoarthritis [114] | Human ESC-MSCs; Tangential flow filtration | 100 μg, three doses (2, 4, and 8-weeks post-injury); intra-articular injection | Not studied | -Suppressed inflammation, apoptosis and matrix degradation -Promoted TMJ repair and regeneration |
| Inflammatory arthritis [115] | Mouse BM-MSC; Differential centrifugation with ultracentrifugation | 250 ng of Exos, 250 or 600 ng of MPs, two doses (Day 18 and post-injury); intravenous | Not studied | -Exerted an anti-inflammatory role on T and B lymphocytes -Exos were more efficient in suppressing inflammation in vivo |
| Articular cartilage [116] | Human ESC-MSCs; Tangential flow filtration | 100 μg, once a week for up to 12 weeks; intra-articular injection | CD73 | -Induced the polarization of M2-like macrophages, and reduced pro-inflammatory cytokine production -Enhanced cellular proliferation and chondrocyte functions |
| Duchenne muscular dystrophy [117] | Placenta-MSC; Differential centrifugation with ultracentrifugation | 5×109 particles, single dose; intra-muscular injection | miR-29c | -Promoted muscle differentiation in vitro -Decreasing inflammation and fibrosis in mdx mice |
| Skeletal muscle damage [118] | Human AD-MSC; Differential centrifugation with ultracentrifugation | total 2×1010 particles: 1×1010 (immediately after injury), intravenous; 0.5×1010 (Day 1 and 2 post-injury), intra-muscular injection | neuregulin 1 protein | -Impaired inflammatory cell infiltration -Induce vascular growth and protect muscle against I/R damage |
| Skeletal muscle damage [119] | Human AD-MSC; Differential centrifugation with ultracentrifugation | 1 μg, single dose; intra-muscular injection | miRNAs | -Triggered macrophage polarization from a M1 to a M2 phenotype -Downregulated the pro-inflammatory cytokine IL-6 accompanied by the upregulation of IL-10 |
| Others | ||||
| Obesity [120] | Mouse AD-MSC; ExoQuick-TC (System Bioscience) | 30 μg, once every 3 days for 6-8 weeks; intraperitoneal | STAT3 protein | -Polarized macrophages toward M2 expressing high levels of arginase-1 and IL-10 -Alleviated white adipose tissue inflammation, obesity, and hepatic steatosis, and improved metabolic homeostasis |
| Inflammatory bowel disease [121] | Human BM-MSC; Differential centrifugation with ultracentrifugation | 200 μg, single dose; intravenous | metallothionein-2 protein | -Downregulated inflammatory responses and maintained intestinal barrier integrity -Polarized M2b macrophages and subsequently induced IL-10 secretion |
| Retinal I/R injury [122] | Human BM-MSC; Ultrafiltration followed by ExoQuick-TC (System Bioscience) | 4×106 particles, single dose; vitreous humor injection | Not studied | -Enhanced functional recovery, and decreased neuro-inflammation and apoptosis |
| Esophageal fistula [123] | Swine AD-MSC; Differential centrifugation with ultracentrifugation | PF-127 gel containing 5.2×1011 particles injected through fistula internal and external orifices, respectively | Not studied | -Reduced the density of myofibroblasts -Decreased fibrosis and inflammatory response -Increased angiogenesis. |
| Burn [124] | Human UC-MSC; PureExo® Exosome Isolation Kit (101Bio) | 800 μg (RNA concentration), single dose; intravenous | miR-181c | -Reduced burn-induced inflammation by downregulating the TLR4 signaling pathway |
AD, adipose tissue; BM, bone marrow; UC, umbilical cord; iPSC, induced pluripotent stem cell; ESC, embryonic stem cell; ILC2, Group 2 innate lymphoid cell; I/R, ischemia/reperfusion; EAE, experimental autoimmune encephalomyelitis; TMJ, temporomandibular joint; Exo, exosome; MP, microparticle; VEGF, vascular endothelial growth factor; KGF, keratinocyte growth factor; MnSOD, manganese Superoxide dismutase.
Concurrently, clinical trials have been initiated to assess the therapeutic value of MSC-EVs. The first documented clinical MSC-EV administration was performed on a steroid-refractory GvHD patient. The therapy reduced the pro-inflammatory cytokine response and attenuated the clinical GvHD symptoms, which were stable even after 4 months following MSC-EV treatment [125].
Subsequently, a randomized, placebo-controlled, phase 2/3 clinical pilot study showed that MSC-EVs safely and efficiently ameliorated the inflammatory immune reaction and improved overall kidney function in grade III-IV CKD patients [126]. In addition, two ongoing trials are executed to use MSC-EVs for the treatment of type I diabetes mellitus (ClinicalTrials.gov identifier: NCT02138331) and macular holes (ClinicalTrials.gov identifier: NCT03437759). These clinical observations, together with evidence from preclinical studies, further indicate that native EVs from MSCs hold therapeutic promise in inflammatory diseases.
However, despite current successes, there are still some hurdles in the therapeutic use of MSC-EVs. First, the composition and function of EVs are susceptible to the producer cell state, and MSCs from different donors add another layer of uncertainty and risk. How to guarantee the stability and security of the MSC-EV therapy is a challenging problem. To address this, the underlying therapeutic mechanisms and pharmacodynamic effects of MSC-EVs should be defined. Second, owing to the lack of comparative studies, it is still unclear whether MSC-EVs work better than their parental cells. MSC-EVs protected against neonatal hyperoxic lung injury as effectively as MSC therapy [96], whereas other studies showed inferior potency of MSC-EVs compared to their parental cells for bone regeneration [127] and atherosclerosis [128]. Furthermore, which subspecies of vesicles are responsible for the therapeutic efficacy remains controversial. A recent study suggested that MSC-derived exosomes were more efficient in suppressing inflammation in treating inflammatory arthritis than microvesicels [115]. Considering the concurrence of pro- and anti-tumorigenic ability of MSC-EVs [129,130], further evaluation of their safety and potential in this regard is needed.
In addition to MSCs, EVs from other sources, such as amniotic epithelial cells [131], endothelial progenitor cells [132,133], and immune cells [134,135], show powerful effects in mitigating inflammation as well. For example, TNF-activated neutrophils released EVs displaying annexin A1 and/or phosphatidylserine on the surface, attenuating inflammation in the synovia by affecting macrophage polarization and potentially also fibroblast-like synoviocytes [136]. Interestingly, microorganism-derived EVs are also considered to be a mine for pharmaceutical exploitation. Akkermansia muciniphila is a beneficial gastrointestinal microbiota whose EVs have recently been introduced to treat high-fat diet-induced obesity and show significant effects on adipose dysfunction, inflammation reduction, and obesity reversal [137].
EVs as nanocarriers for anti-inflammatory therapeutics
Respiratory system inflammation
Acute respiratory distress syndrome (ARDS), a severe progression of acute lung injury (ALI), seriously threatens the lives of intensive care unit patients. However, there is no definitive pharmacotherapy in place for ARDS/ALI patients, in part because drugs cannot efficiently target inflamed lungs. Gao et al. reported a strategy that applied neutrophil-derived nitrogen cavitation (a physical force)-induced EVs to deliver the anti-inflammatory drug piceatannol (Pic-NC-EVs) for ALI treatment. As a result, Pic-NC-EVs significantly attenuated acute lung inflammation by inhibiting the NF-κB pathway and thus prolonged the mouse survival in sepsis [49]. Moreover, Zhang et al. delivered anti-inflammatory small RNA molecules, such as Myd88 siRNA, miR-15a mimic, or miR-155 inhibitor, into lung macrophages respectively using serum-derived EVs as vehicles, successfully relieving LPS-induced lung inflammation [138].
Asthma is a common chronic respiratory disease characterized by airway remodeling and airway inflammation. Anti-inflammatory and anti-airway remodeling therapies are the two basic treatments. Recently, Shang et al. modified MSCs to generate EVs enriched with the circular RNA-mmu_circ_0001359, which enhanced FoxO1 signaling-mediated M2-like macrophage activation via sponging miR-183-5p and thereby suppressing pro-inflammatory cytokine expression and attenuating airway remodeling [139]. The findings above highlight the promise of EV-based nanotherapeutics for lung inflammation. Furthermore, compared to artificial nanoparticles, EVs may not induce cytotoxicity, inflammation, changes in airway resistance or structural damage of the lung tissue.
Cardiovascular disease
Cardiovascular disease (CVD) is the leading cause of morbidity and mortality worldwide. Inflammatory processes are firmly established as central to the development and complications of CVD and have become a target of therapy. Increasing evidence has shown that EV-based therapy represents a promising tool to combat inflammation in CVD, such as myocardial infarction, stroke, pulmonary hypertension and others [140]. For example, localized injection of miRNA-181a- overexpressing MSC-EVs inhibited the inflammatory response and raised the Treg cell ratio through inhibition of c-Fos protein, encouraging restoration of the infarcted hearts [141]. However, the injected EVs may be eliminated quickly due to a lack of support, since they were undetectable 3 h after myocardial injection [142]. To prevent this rapid elimination, MSC-EVs were encapsulated in functional peptide hydrogels to prolong EV retention in the heart, which resulted in better cardiac function with reduced apoptosis, inflammation and fibrosis after myocardial infarction when compared with free EVs [143]. Of note, it remains to be established whether the systemic delivery of EVs would trigger similar positive effects because circulating EVs may be rapidly cleared by the mononuclear phagocyte system, and the continuous shear stress from constant blood flow may prevent nontargeted EVs from accumulating in the heart.
Neuroinflammation
EV-based nanotherapeutics have been widely applied in the treatment of nervous system diseases, such as Alzheimer's disease [24], Huntington's disease [71,144], Parkinson's disease [145-148], and ischemic reperfusion injury [149,150]. For example, Parkinson's disease is a neurodegenerative disorder marked by the elevation of α-synuclein, brain inflammation, and reactive oxygen species. Haney et al. harnessed EV therapy for the intranasal delivery of catalase to the brain, reducing brain oxidative stress and inflammation and improving neuronal survival in a mouse model of Parkinson's disease [145]. Kalani et al. proved the neuroprotective effects, such as reduced inflammation, restored blood brain barrier and overall improved neurological score, of curcumin-loaded embryonic stem cell-derived EVs in an ischemia-reperfusion injury model [149]. Dong et al. reported that neutrophil membrane-derived vesicles can specifically target inflamed brain endothelium during ischemia-reperfusion injury and deliver RvD2 to prevent neuroinflammation [50]. Importantly, these studies suggest that the nanosize, cellular origin, and other properties made EVs capable of passing through the blood brain barrier, thus highlighting the potential of EV-based nanotherapeutics for the treatment of central nervous system disorders.
Autoimmune diseases
Autoimmune diseases refer to a group of chronic inflammatory diseases caused by self-immune responses to autoantigens; such conditions include rheumatoid arthritis, systemic lupus erythematosus, type 1 diabetes, and multiple sclerosis, among others. The current conventional therapies, notably corticosteroids and immunosuppressors, seriously endanger human health. To solve this dilemma, EV-based nanotherapeutics have been tested, and certain results have been achieved in treating rheumatoid arthritis [151] and multiple sclerosis [44,91,152]. Chen et al. transfected MSCs with a miR-150-5p expression plasmid to harvest miR-150-5p-enriched EVs, which significantly reduced hind paw thickness and clinical arthritic scores in collagen-induced arthritis mice by inhibiting synoviocyte hyperplasia and angiogenesis [151]. Yu et al. generated therapeutic EVs from modified dendritic cells expressing membrane‐associated TGF‐β1 (mTGF‐β1). The mTGF‐β1-EVs possessed powerful immunosuppressive ability and effectively inhibited the development and progression of multiple sclerosis in different strains of mice [152]. Moreover, the aforementioned EVs loaded with the anti-inflammatory drug curcumin [44] or IL-4 protein [91] have also been proven to reduce clinical signs in a mouse model of multiple sclerosis.
Renal inflammation
The role of EVs in the pathogenesis of renal inflammation is widely accepted [153-155], but their therapeutic application has just begun. In the first study, MSC-EVs were used to deliver miR-let7c to damaged kidneys, achieving the repression of renal fibrosis by down-regulating TGF-β1 protein expression in UUO mice [156]. A subsequent study employed macrophage-derived EVs for the targeted delivery of dexamethasone to the inflamed endothelium, not only improving the anti-inflammatory efficacy but also reducing the side effects of dexamethasone. Interestingly, in addition to the drug, glucocorticoid receptors in the macrophages were also packaged and delivered to renal cells, thereby increasing cellular levels of the receptor and improving cell sensitivity to dexamethasone [46]. Considering that the downregulation of glucocorticoid receptors is associated with acquired resistance [157], such EVs loaded with the drug and adjunct receptors may be beneficial for steroid-resistant patients. Furthermore, these findings provide some preliminary evidence that in the context of kidney injury, EVs can cross the glomerular filtration barrier by right of their small size and good biocompatibility; however, additional discussion and analyses are needed to clarify this issue.
Other diseases
Inflammatory bowel disease, including Crohn's disease and ulcerative colitis, is a chronic and relapsing inflammatory condition of the gastrointestinal tract. Current medical management strategies cannot keep patients in remission for the long term or may cause serious side effects due to the typically high dosage required. Recently, the well-known anti-inflammatory miR-146a was packaged into MSC-EVs for the treatment of experimental colitis, leading to inhibition of the NF-κB-dependent activation of inflammatory gene transcription by targeting TNF receptor-associated factor 6 and IL-1 receptor-associated kinase 1 in colon tissue [158].
Moreover, EV-based nanotherapeutics have also been applied in a murine model of duchenne muscular dystrophy [72,159], a fatal neuromuscular disorder that manifests as progressive muscle wasting. For instance, myostatin propeptide, which is a natural inhibitor of mature myostatin, was anchored to the surface of NIH3T3 mouse embryonic fibroblast cell-derived EVs by fusing the inhibitory domain of myostatin propeptide into the extracellular loop of CD63, which enhanced the serum stability and delivery efficiency of the propeptide and thereby led to a significant increase in muscle mass and functional rescue in mdx mice [159].
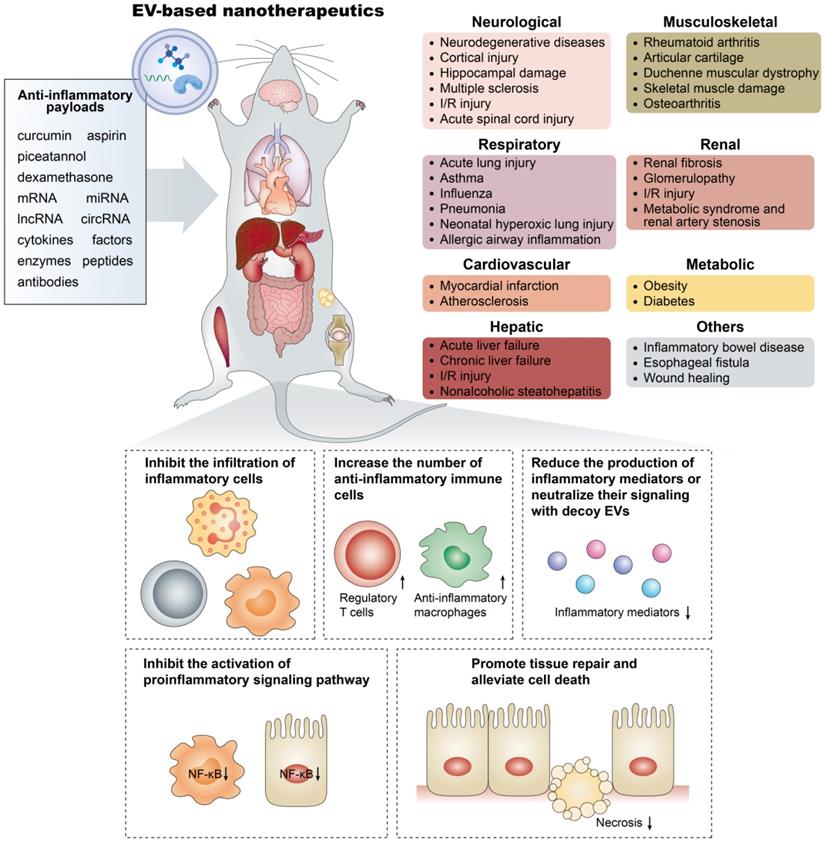
Overall, these examples underline the versatility of EV-based nanotherapeutics and their potential for addressing multiple unmet clinical demands (Figure 3). However, in addition to their efficacy, researchers should tend to attach great importance to the pharmacokinetic and pharmacodynamic profiles of EV-based nanotherapeutics to optimize and standardize their treatment strategies to hasten their clinical development. It should be noted that due to the length limitations, this review focuses on the anti-inflammatory applications of EVs in noncancer diseases, whereas comprehensive reviews on their roles in anticancer immunity are available elsewhere [160,161].
Emerging EV-based nanotherapeutics in the treatment of inflammatory-related diseases. The anti-inflammatory payloads in EV-based nanotherapeutics include small molecule drugs, RNA species, and therapeutic proteins, which show significant therapeutic efficacy in inflammatory-related diseases occurring in organs such as the brain, lung, heart, liver, and kidney through various mechanisms.
Challenges and perspectives in the anti-inflammatory application of EV-based nanotherapeutics
EV-based nanotherapeutics represent a promising next-generation treatment for many inflammatory diseases by virtue of their own merits, such as desirable stability in body fluids, ability to surmount biological barriers, low immunogenicity when using autologously derived EVs, fitness for the delivery of various therapeutic agents, and ability to achieve disease-specific tailor-made delivery, which can provide many dramatic improvements to an army of drugs to fight excessive or persistent inflammation. However, a set of common concerns in the process of EV production, functionalization, and application should be ironed out before turning them into the clinic (Figure 4):
The flow and common concerns of developing EV-based nanotherapeutics.
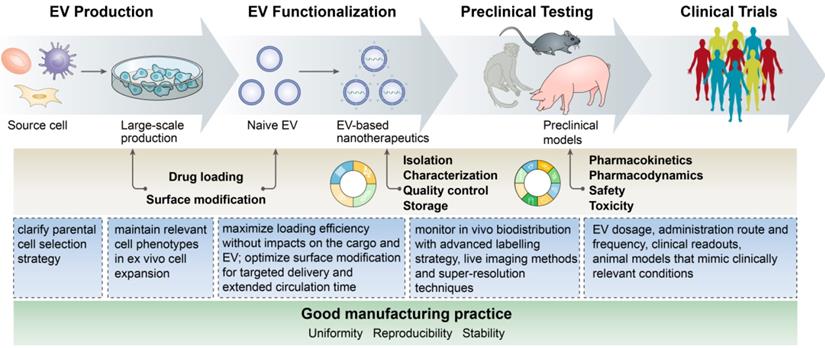
- Source cell selection: Almost all cell types can secrete EVs, but which source is suitable for EV production for anti-inflammatory applications remains unclear. Autologous primary cell-derived EVs have a low risk of immunological rejection, but suffer from lower yield, whereas cell lines can be cultured indefinitely but may carry some risk of oncogenic potential. Despite success in developing immune cell-derived EV-based delivery system, however, it should be noted that these EVs may also have the potential to provoke inflammation. How the EV source affects therapeutic efficacy and immune response needs further investigation.
- Unwanted biological effects of the inherent cargos in EVs: The capacity of EVs for the intracellular transmission of bioactive molecules influences the physiological and pathological functions of recipient cells. For example, heat shock proteins present on EVs may trigger unintentional immune cell activation, and consequently impact therapeutic efficacy, which deserves serious consideration when designing EV-based therapeutics. An important question is how to selectively dislodge undesirable substances from EVs during biogenesis.
- Large-scale production: Developing EV-based nanotherapeutics requires a production method that assures not only high quality but also high quantity. However, existing methods cannot, as yet, fully meet the standards for clinical translation. Emerging cell culture equipment, such as a hollow-fiber culture system, has been generated for the scalable and continuous production of therapeutic EVs, affording 40-fold more EVs per volume of conditioned medium than conventional cell culture [162]. Moreover, physical and mechanical stress, such as sonication, extrusion and centrifugal force, have been utilized to yield EV-like cell-derived vesicles, increasing EV production by more than 100-fold compared to naturally secreted EVs [40,163,164]. However, these EV mimetics may not necessarily share all common features with biogenic EVs, and whether this impacts delivery, biocompatibility, immunogenicity, or other EV properties will require stringent assessment.
- Optimal cargo loading: There is no doubt that the loading efficiency of EVs for different types of cargos largely determines the success or failure of EV-based therapy. Excessively rough loading approaches may ruin the EV integrity, induce immunogenicity, or lead to drug degradation and EV aggregation. Procedures that maximize the loading efficiency without compromising the stability of the cargo or the EVs themselves should be defined. For the endogenous loading, the mechanisms that mediate the sorting of cargos into EVs remain to be fully unraveled.
- Nonspecific uptake by the mononuclear phagocyte system: The major limitation of EV delivery is its inability to reach disease sites owing to nonspecific uptake by the mononuclear phagocyte system. Recently, Wan et al. reported that prior blockade of the mononuclear phagocyte system with siCltc-modified EVs significantly improved the protective effect of the miRNA-21-loaded EVs in doxorubicin-induced myocardial toxicity [165], suggesting that the two-step EV delivery strategy (blocking the uptake of EVs before the delivery of therapeutic EVs) would be a promising method for the treatment of heart diseases.
- Monitoring the in vivo biodistribution: Labeling strategies such as fluorescent dyes and luciferase have been used for the in vivo tracing of EVs [166]. Direct labeling with fluorescent dyes is easy but may affect the biological behavior of the EVs. Leftover dyes could be detected rather than vesicles and may increase the risk of tissue contamination. In addition, current techniques cannot measure the precise amount of EVs in specific organs due to the thickness of the tissue and the resulting loss of signal. Thus, advances in labeling strategies, in vivo imaging techniques and super-resolution construction will surely aid in the dynamic tracking of EVs and their intracellular fates.
- In vivo assays and safety challenges: Although trials are already investigating the clinical potential of EV-based therapy, there is still a lack of the application note regarding many factors, such as EV dosage, administration route and frequency, and clinical readouts. Animal models that mimic clinically relevant conditions are urgently needed to evaluate the safety, toxicity and pharmacokinetics of EV-based nanotherapeutics to support clinical application.
Moreover, to broaden the application of EVs in the inflammatory diseases, several issues need to be considered:
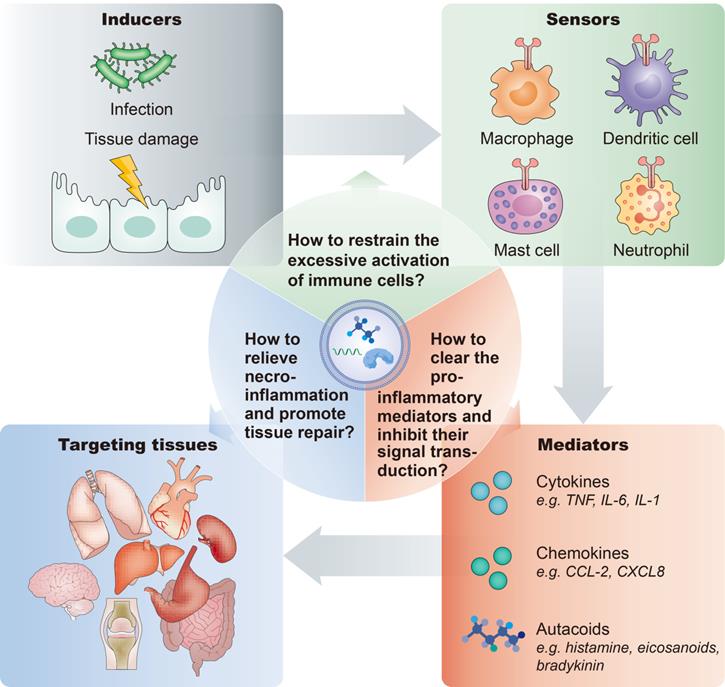
- First, the inflammatory response is orchestrated by various modulators and pathways. In general, a typical inflammatory pathway consists of inducers, sensors, mediators and effectors. Exogenous or endogenous inducers of inflammation initiate and activate the sensors, such as pattern recognition receptors expressed on immune cells, which instigate the production and secretion of various inflammatory mediators, such as cytokines, chemokines, and autacoids, thereby affecting the functions of target organs (Figure 5). Thus, developing rational targeting or therapeutic strategies tailored to these different stages will advance the application of EV-based nanotherapeutics for multiple inflammatory-related diseases.
- Second, it is important to take advantage of natural EVs with innate therapeutic benefit. For example, stem cell-derived EVs exhibit versatile functions, including anti-apoptosis, anti-inflammation, and anti-fibrosis activities. Exploiting these EVs as delivery vectors for anti-inflammatory agents may provide synergy and facilitate the treatment effect. Meanwhile, they may also offset the side effects of packaged drugs, such as aspirin, which can markedly inhibit inflammation but induce apoptosis.
- Additionally, the application of EVs in the anti-inflammatory therapy is still in the stage of preliminary development, and further exploitation and utilization are needed to maximize their advantages to develop theranostic platforms for anti-inflammatory therapy. For example, EVs in the circulation and body fluid can provide diagnostic information and aid in making therapeutic decisions for inflammatory disorders. EVs combined with imaging modalities will enable the visualization of inflamed tissues, enabling noninvasive monitoring of the in vivo efficacy of an anti-inflammatory drug.
Inflammatory pathway components and key considerations of anti-inflammatory treatment. A typical inflammatory pathway consists of inducers, sensors, mediators and effectors. For the anti-inflammatory therapy, important questions are: How to restrain the excessive activation of immune cells? How to clear the pro-inflammatory mediators and inhibit their signal transduction? How to inhibit necroinflammation and promote tissue repair?
Conclusions
The past few decades have provided a wealth of information on how maladaptive and excessive inflammation drives a number of diseases. However, translating this knowledge to improve clinical therapy is confronted with many challenges. The field of EV-based nanotherapeutics is rapidly evolving and expanding, opening up vast opportunities for a new generation of anti-inflammatory medicines. Two overall applications are emerging: (i) exploiting the intrinsic therapeutic activity of EVs for inflammation-combating; and (ii) employing EVs as nature's own delivery carrier for small molecule drugs, therapeutic RNAs and proteins delivery in combination with targeting moieties. Despite the inspiring therapeutic potential, the field is being starved of new techniques to optimize EV production and cargo loading, target single-vesicle analysis, and track EVs with powerful imaging methods. In the future, deeper and broader cooperation in areas of medicine, nanotechnology, material science, bioengineering and pharmaceutical perspectives will surely enable better use of EVs in the treatment of inflammation.
Abbreviations
EVs: extracellular vesicles; PEG: polyethylene glycol; ICAM-1: intercellular cell adhesion molecule-1; VCAM-1: vascular cell adhesion molecule-1; RVG: rabies viral glycoprotein; RGD: Arg-Gly-Asp; RGE: neuropilin-1-targeted peptide (RGERPPR); EXPLORs: exosomes for protein loading via optically reversible protein-protein interactions; RNAi: RNA interference; EXOtic devices: EXOsomal transfer into cells devices; MSC: mesenchymal stem cells; IL-1RA: interleukin-1 receptor antagonist; ARDS: acute respiratory distress syndrome; CVD: cardiovascular disease; UUO: unilateral ureteral occlusion.
Acknowledgements
This study was supported by grants from the National Natural Science Foundation of China (No. 81720108007, 81670696, 81470922 and 31671194), the National Key Research and Development Program (2018YFC1314000), the Clinic Research Center of Jiangsu Province (No. BL2014080), and the Postgraduate Research & Practice Innovation Program of Jiangsu Province (No. KYCX18_0171).
Competing Interests
The authors have declared that no competing interest exists.
References
1. Medzhitov R. Origin and physiological roles of inflammation. Nature. 2008;454:428-35
2. Medzhitov R. Inflammation 2010: new adventures of an old flame. Cell. 2010;140:771-6
3. Kotas ME, Medzhitov R. Homeostasis, inflammation, and disease susceptibility. Cell. 2015;160:816-27
4. Karin M, Clevers H. Reparative inflammation takes charge of tissue regeneration. Nature. 2016;529:307-15
5. GBD 2017 Causes of Death Collaborators. Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980-2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet. 2018;392:1736-88
6. Bonaccio M, Di Castelnuovo A, Pounis G, De Curtis A, Costanzo S, Persichillo M. et al. A score of low-grade inflammation and risk of mortality: prospective findings from the Moli-sani study. Haematologica. 2016;101:1434-41
7. Hanke T, Merk D, Steinhilber D, Geisslinger G, Schubert-Zsilavecz M. Small molecules with anti-inflammatory properties in clinical development. Pharmacol Ther. 2016;157:163-87
8. Tabas I, Glass CK. Anti-inflammatory therapy in chronic disease: challenges and opportunities. Science. 2013;339:166-72
9. Alaarg A, Pérez-Medina C, Metselaar JM, Nahrendorf M, Fayad ZA, Storm G. et al. Applying nanomedicine in maladaptive inflammation and angiogenesis. Adv Drug Deliv Rev. 2017;119:143-58
10. EL Andaloussi S, Mäger I, Breakefield XO, Wood MJ. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat Rev Drug Discov. 2013;12:347-57
11. Agrahari V, Agrahari V, Burnouf PA, Chew CH, Burnouf T. Extracellular microvesicles as new industrial therapeutic frontiers. Trends Biotechnol. 2019;37:707-29
12. Wiklander OPB, Brennan MÁ, Lötvall J, Breakefield XO, El Andaloussi S. Advances in therapeutic applications of extracellular vesicles. Sci Transl Med. 2019;11:eaav8521
13. Liu C, Su C. Design strategies and application progress of therapeutic exosomes. Theranostics. 2019;9:1015-28
14. Tang TT, Lv LL, Lan HY, Liu BC. Extracellular vesicles: opportunities and challenges for the treatment of renal diseases. Front Physiol. 2019;10:226
15. Gao J, Dong X, Wang Z. Generation, purification and engineering of extracellular vesicles and their biomedical applications. Methods. 2020;177:114-25
16. van Niel G, D'Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19:213-28
17. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367:eaau6977
18. Colombo M, Raposo G, Théry C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30:255-89
19. Tkach M, Théry C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell. 2016;164:1226-32
20. Mathieu M, Martin-Jaular L, Lavieu G, Théry C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019;21:9-17
21. Lv LL, Feng Y, Tang TT, Liu BC. New insight into the role of extracellular vesicles in kidney disease. J Cell Mol Med. 2019;23:731-9
22. Piffoux M, Nicolás-Boluda A, Mulens-Arias V, Richard S, Rahmi G, Gazeau F. et al. Extracellular vesicles for personalized medicine: The input of physically triggered production, loading and theranostic properties. Adv Drug Deliv Rev. 2019;138:247-58
23. Gudbergsson JM, Jønsson K, Simonsen JB, Johnsen KB. Systematic review of targeted extracellular vesicles for drug delivery-Considerations on methodological and biological heterogeneity. J Control Release. 2019;306:108-20
24. Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJ. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol. 2011;29:341-5
25. Wiklander OP, Nordin JZ, O'Loughlin A, Gustafsson Y, Corso G, Mäger I. et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J Extracell Vesicles. 2015;4:26316
26. Tian Y, Li S, Song J, Ji T, Zhu M, Anderson GJ. et al. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials. 2014;35:2383-90
27. Hung ME, Leonard JN. Stabilization of exosome-targeting peptides via engineered glycosylation. J Biol Chem. 2015;290:8166-72
28. Ohno S, Takanashi M, Sudo K, Ueda S, Ishikawa A, Matsuyama N. et al. Systemically injected exosomes targeted to EGFR deliver antitumor microRNA to breast cancer cells. Mol Ther. 2013;21:185-91
29. Kooijmans SA, Aleza CG, Roffler SR, van Solinge WW, Vader P, Schiffelers RM. Display of GPI-anchored anti-EGFR nanobodies on extracellular vesicles promotes tumour cell targeting. J Extracell Vesicles. 2016;5:31053
30. Longatti A, Schindler C, Collinson A, Jenkinson L, Matthews C, Fitzpatrick L. et al. High affinity single-chain variable fragments are specific and versatile targeting motifs for extracellular vesicles. Nanoscale. 2018;10:14230-44
31. Wang JH, Forterre AV, Zhao J, Frimannsson DO, Delcayre A, Antes TJ. et al. Anti-HER2 scFv-directed extracellular vesicle-mediated mRNA-based gene delivery inhibits growth of HER2-positive human breast tumor xenografts by prodrug activation. Mol Cancer Ther. 2018;17:1133-42
32. Yang Y, Hong Y, Nam GH, Chung JH, Koh E, Kim IS. Virus-mimetic fusogenic exosomes for direct delivery of integral membrane proteins to target cell membranes. Adv Mater. 2017;29:1605604
33. Kamerkar S, LeBleu VS, Sugimoto H, Yang S, Ruivo CF, Melo SA. et al. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature. 2017;546:498-503
34. Presolski SI, Hong VP, Finn MG. Copper-catalyzed azide-alkyne click chemistry for bioconjugation. Curr Protoc Chem Biol. 2011;3:153-62
35. Smyth T, Petrova K, Payton NM, Persaud I, Redzic JS, Graner MW. et al. Surface functionalization of exosomes using click chemistry. Bioconjug Chem. 2014;25:1777-84
36. Tian T, Zhang HX, He CP, Fan S, Zhu YL, Qi C. et al. Surface functionalized exosomes as targeted drug delivery vehicles for cerebral ischemia therapy. Biomaterials. 2018;150:137-49
37. Jia G, Han Y, An Y, Ding Y, He C, Wang X. et al. NRP-1 targeted and cargo-loaded exosomes facilitate simultaneous imaging and therapy of glioma in vitro and in vivo. Biomaterials. 2018;178:302-16
38. Sato YT, Umezaki K, Sawada S, Mukai SA, Sasaki Y, Harada N. et al. Engineering hybrid exosomes by membrane fusion with liposomes. Sci Rep. 2016;6:21933
39. Piffoux M, Silva AKA, Wilhelm C, Gazeau F, Tareste D. Modification of extracellular vesicles by fusion with liposomes for the design of personalized biogenic drug delivery systems. ACS Nano. 2018;12:6830-42
40. Wan Y, Wang L, Zhu C, Zheng Q, Wang G, Tong J. et al. Aptamer-conjugated extracellular nanovesicles for targeted drug delivery. Cancer Res. 2018;78:798-808
41. Pi F, Binzel DW, Lee TJ, Li Z, Sun M, Rychahou P. et al. Nanoparticle orientation to control RNA loading and ligand display on extracellular vesicles for cancer regression. Nat Nanotechnol. 2018;13:82-9
42. Kooijmans SAA, Fliervoet LAL, van der Meel R, Fens MHAM, Heijnen HFG, van Bergen En Henegouwen PMP. et al. PEGylated and targeted extracellular vesicles display enhanced cell specificity and circulation time. J Control Release. 2016;224:77-85
43. Nakase I, Futaki S. Combined treatment with a pH-sensitive fusogenic peptide and cationic lipids achieves enhanced cytosolic delivery of exosomes. Sci Rep. 2015;5:10112
44. Zhuang X, Xiang X, Grizzle W, Sun D, Zhang S, Axtell RC. et al. Treatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brain. Mol Ther. 2011;19:1769-79
45. Nolte-'t Hoen EN, Buschow SI, Anderton SM, Stoorvogel W, Wauben MH. Activated T cells recruit exosomes secreted by dendritic cells via LFA-1. Blood. 2009;113:1977-81
46. Tang TT, Lv LL, Wang B, Cao JY, Feng Y, Li ZL. et al. Employing macrophage-derived microvesicle for kidney-targeted delivery of dexamethasone: An efficient therapeutic strategy against renal inflammation and fibrosis. Theranostics. 2019;9:4740-55
47. Yuan D, Zhao Y, Banks WA, Bullock KM, Haney M, Batrakova E. et al. Macrophage exosomes as natural nanocarriers for protein delivery to inflamed brain. Biomaterials. 2017;142:1-12
48. Gao J, Chu D, Wang Z. Cell membrane-formed nanovesicles for disease-targeted delivery. J Control Release. 2016;224:208-16
49. Gao J, Wang S, Wang Z. High yield, scalable and remotely drug-loaded neutrophil-derived extracellular vesicles (EVs) for anti-inflammation therapy. Biomaterials. 2017;135:62-73
50. Dong X, Gao J, Zhang CY, Hayworth C, Frank M, Wang Z. Neutrophil membrane-derived nanovesicles alleviate inflammation to protect mouse brain injury from ischemic stroke. ACS Nano. 2019;13:1272-83
51. Chu D, Dong X, Shi X, Zhang C, Wang Z. Neutrophil-based drug delivery systems. Adv Mater. 2018;30:e1706245
52. Ley K, Laudanna C, Cybulsky MI, Nourshargh S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nat Rev Immunol. 2007;7:678-89
53. Buschow SI, Nolte-'t Hoen EN, van Niel G, Pols MS, ten Broeke T, Lauwen M. et al. MHC II in dendritic cells is targeted to lysosomes or T cell-induced exosomes via distinct multivesicular body pathways. Traffic. 2009;10:1528-42
54. Denzer K, van Eijk M, Kleijmeer MJ, Jakobson E, de Groot C, Geuze HJ. Follicular dendritic cells carry MHC class II-expressing microvesicles at their surface. J Immunol. 2000;165:1259-65
55. Cheng Q, Shi X, Han M, Smbatyan G, Lenz HJ, Zhang Y. Reprogramming exosomes as nanoscale controllers of cellular immunity. J Am Chem Soc. 2018;140:16413-17
56. Wang X, Chen Y, Zhao Z, Meng Q, Yu Y, Sun J. et al. Engineered exosomes with ischemic myocardium-targeting peptide for targeted therapy in myocardial infarction. J Am Heart Assoc. 2018;7:e008737
57. Kim H, Yun N, Mun D, Kang JY, Lee SH, Park H. et al. Cardiac-specific delivery by cardiac tissue-targeting peptide-expressing exosomes. Biochem Biophys Res Commun. 2018;499:803-8
58. Zhang N, Chittasupho C, Duangrat C, Siahaan TJ, Berkland C. PLGA nanoparticle-peptide conjugate effectively targets intercellular cell-adhesion molecule-1. Bioconjug Chem. 2008;19:145-52
59. Chittasupho C, Xie SX, Baoum A, Yakovleva T, Siahaan TJ, Berkland CJ. ICAM-1 targeting of doxorubicin-loaded PLGA nanoparticles to lung epithelial cells. Eur J Pharm Sci. 2009;37:141-50
60. Nahrendorf M, Jaffer FA, Kelly KA, Sosnovik DE, Aikawa E, Libby P. et al. Noninvasive vascular cell adhesion molecule-1 imaging identifies inflammatory activation of cells in atherosclerosis. Circulation. 2006;114:1504-11
61. Zhao Z, Ukidve A, Kim J, Mitragotri S. Targeting Strategies for Tissue-Specific Drug Delivery. Cell. 2020;181:151-67
62. Salazar MD, Ratnam M. The folate receptor: what does it promise in tissue-targeted therapeutics? Cancer Metastasis Rev. 2007;26:141-52
63. Tran PH, Xiang D, Nguyen TN, Tran TT, Chen Q, Yin W. et al. Aptamer-guided extracellular vesicle theranostics in oncology. Theranostics. 2020;10:3849-66
64. Li C, Guo F, Wang X, Liu D, Wu B, Wang F. et al. Exosome-based targeted RNA delivery for immune tolerance induction in skin transplantation. J Biomed Mater Res A. 2020;108:1493-500
65. Zhang W, Yu ZL, Wu M, Ren JG, Xia HF, Sa GL. et al. Magnetic and folate functionalization enables rapid isolation and enhanced tumor-targeting of cell-derived microvesicles. ACS Nano. 2017;11:277-90
66. Sun D, Zhuang X, Xiang X, Liu Y, Zhang S, Liu C. et al. A novel nanoparticle drug delivery system: the anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomes. Mol Ther. 2010;18:1606-14
67. Fuhrmann G, Serio A, Mazo M, Nair R, Stevens MM. Active loading into extracellular vesicles significantly improves the cellular uptake and photodynamic effect of porphyrins. J Control Release. 2015;205:35-44
68. Shtam TA, Kovalev RA, Varfolomeeva EY, Makarov EM, Kil YV, Filatov MV. Exosomes are natural carriers of exogenous siRNA to human cells in vitro. Cell Commun Signal. 2013;11:88
69. Kooijmans SAA, Stremersch S, Braeckmans K, de Smedt SC, Hendrix A, Wood MJA. et al. Electroporation-induced siRNA precipitation obscures the efficiency of siRNA loading into extracellular vesicles. J Control Release. 2013;172:229-38
70. Haraszti RA, Miller R, Didiot MC, Biscans A, Alterman JF, Hassler MR. et al. Optimized cholesterol-siRNA chemistry improves productive loading onto extracellular vesicles. Mol Ther. 2018;26:1973-82
71. Didiot MC, Hall LM, Coles AH, Haraszti RA, Godinho BM, Chase K. et al. Exosome-mediated delivery of hydrophobically modified siRNA for huntingtin mRNA silencing. Mol Ther. 2016;24:1836-47
72. Gao X, Ran N, Dong X, Zuo B, Yang R, Zhou Q. et al. Anchor peptide captures, targets, and loads exosomes of diverse origins for diagnostics and therapy. Sci Transl Med. 2018;10:eaat0195
73. Villarroya-Beltri C, Gutiérrez-Vázquez C, Sánchez-Cabo F, Pérez-Hernández D, Vázquez J, Martin-Cofreces N. et al. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat Commun. 2013;4:2980
74. Lee H, Li C, Zhang Y, Zhang D, Otterbein LE, Jin Y. Caveolin-1 selectively regulates microRNA sorting into microvesicles after noxious stimuli. J Exp Med. 2019;216:2202-20
75. Shurtleff MJ, Temoche-Diaz MM, Karfilis KV, Ri S, Schekman R. Y-box protein 1 is required to sort microRNAs into exosomes in cells and in a cell-free reaction. Elife. 2016;5:e19276
76. Santangelo L, Giurato G, Cicchini C, Montaldo C, Mancone C, Tarallo R. et al. The RNA-binding protein SYNCRIP is a component of the hepatocyte exosomal machinery controlling microRNA sorting. Cell Rep. 2016;17:799-808
77. Li Z, Zhou X, Wei M, Gao X, Zhao L, Shi R. et al. In vitro and in vivo RNA inhibition by CD9-HuR functionalized exosomes encapsulated with miRNA or CRISPR/dCas9. Nano Lett. 2019;19:19-28
78. Yim N, Ryu SW, Choi K, Lee KR, Lee S, Choi H. et al. Exosome engineering for efficient intracellular delivery of soluble proteins using optically reversible protein-protein interaction module. Nat Commun. 2016;7:12277
79. Yang Z, Shi J, Xie J, Wang Y, Sun J, Liu T. et al. Large-scale generation of functional mRNA-encapsulating exosomes via cellular nanoporation. Nat Biomed Eng. 2020;4:69-83
80. Kalinec GM, Gao L, Cohn W, Whitelegge JP, Faull KF, Kalinec F. Extracellular vesicles from auditory cells as nanocarriers for anti-inflammatory drugs and pro-resolving mediators. Front Cell Neurosci. 2019;13:530
81. Momen-Heravi F, Bala S, Bukong T, Szabo G. Exosome-mediated delivery of functionally active miRNA-155 inhibitor to macrophages. Nanomedicine. 2014;10:1517-27
82. Gebert LFR, MacRae IJ. Regulation of microRNA function in animals. Nat Rev Mol Cell Biol. 2019;20:21-37
83. Walker S, Busatto S, Pham A, Tian M, Suh A, Carson K. et al. Extracellular vesicle-based drug delivery systems for cancer treatment. Theranostics. 2019;9:8001-17
84. Jiang L, Gu Y, Du Y, Liu J. Exosomes: diagnostic biomarkers and therapeutic delivery vehicles for cancer. Mol Pharm. 2019;16:3333-49
85. Usman WM, Pham TC, Kwok YY, Vu LT, Ma V, Peng B. et al. Efficient RNA drug delivery using red blood cell extracellular vesicles. Nat Commun. 2018;9:2359
86. Wahlgren J, De L Karlson T, Brisslert M, Vaziri Sani F, Telemo E, Sunnerhagen P. et al. Plasma exosomes can deliver exogenous short interfering RNA to monocytes and lymphocytes. Nucleic Acids Res. 2012;40:e130
87. El Andaloussi S, Lakhal S, Mäger I, Wood MJ. Exosomes for targeted siRNA delivery across biological barriers. Adv Drug Deliv Rev. 2013;65:391-7
88. Kojima R, Bojar D, Rizzi G, Hamri GC, El-Baba MD, Saxena P. et al. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson's disease treatment. Nat Commun. 2018;9:1305
89. Leader B, Baca QJ, Golan DE. Protein therapeutics: a summary and pharmacological classification. Nat Rev Drug Discov. 2008;7:21-39
90. Kou X, Xu X, Chen C, Sanmillan ML, Cai T, Zhou Y, Giraudo C. et al. The Fas/Fap-1/Cav-1 complex regulates IL-1RA secretion in mesenchymal stem cells to accelerate wound healing. Sci Transl Med. 2018;10:eaai8524
91. Casella G, Colombo F, Finardi A, Descamps H, Ill-Raga G, Spinelli A. et al. Extracellular vesicles containing IL-4 modulate neuroinflammation in a mouse model of multiple sclerosis. Mol Ther. 2018;26:2107-18
92. Choi H, Kim Y, Mirzaaghasi A, Heo J, Kim YN, Shin JH. et al. Exosome-based delivery of super-repressor IκBα relieves sepsis-associated organ damage and mortality. Sci Adv. 2020;6:eaaz6980
93. Ouyang W, O'Garra A. IL-10 family cytokines IL-10 and IL-22: from basic science to clinical translation. Immunity. 2019;50:871-91
94. Duong N, Curley K, Brown A, Campanelli A, Do MA, Levy D. et al. Decoy exosomes as a novel biologic reagent to antagonize inflammation. Int J Nanomedicine. 2019;14:3413-25
95. Khatri M, Richardson LA, Meulia T. Mesenchymal stem cell-derived extracellular vesicles attenuate influenza virus-induced acute lung injury in a pig model. Stem Cell Res Ther. 2018;9:17
96. Ahn SY, Park WS, Kim YE, Sung DK, Sung SI, Ahn JY. et al. Vascular endothelial growth factor mediates the therapeutic efficacy of mesenchymal stem cell-derived extracellular vesicles against neonatal hyperoxic lung injury. Exp Mol Med. 2018;50:26
97. Monsel A, Zhu YG, Gennai S, Hao Q, Hu S, Rouby JJ. et al. Therapeutic effects of human mesenchymal stem cell-derived microvesicles in severe pneumonia in mice. Am J Respir Crit Care Med. 2015;192:324-36
98. Fang SB, Zhang HY, Wang C, He BX, Liu XQ, Meng XC. et al. Small extracellular vesicles derived from human mesenchymal stromal cells prevent group 2 innate lymphoid cell-dominant allergic airway inflammation through delivery of miR-146a-5p. J Extracell Vesicles. 2020;9:1723260
99. Zhao J, Li X, Hu J, Chen F, Qiao S, Sun X. et al. Mesenchymal stromal cell-derived exosomes attenuate myocardial ischaemia-reperfusion injury through miR-182-regulated macrophage polarization. Cardiovasc Res. 2019;115:1205-16
100. Teng X, Chen L, Chen W, Yang J, Yang Z, Shen Z. Mesenchymal stem cell-derived exosomes improve the microenvironment of infarcted myocardium contributing to angiogenesis and anti-inflammation. Cell Physiol Biochem. 2015;37:2415-24
101. Wang X, Gu H, Qin D, Yang L, Huang W, Essandoh K. et al. Exosomal miR-223 contributes to mesenchymal stem cell-elicited cardioprotection in polymicrobial sepsis. Sci Rep. 2015;5:13721
102. Yao J, Zheng J, Cai J, Zeng K, Zhou C, Zhang J. et al. Extracellular vesicles derived from human umbilical cord mesenchymal stem cells alleviate rat hepatic ischemia-reperfusion injury by suppressing oxidative stress and neutrophil inflammatory response. FASEB J. 2019;33:1695-710
103. Mardpour S, Ghanian MH, Sadeghi-Abandansari H, Mardpour S, Nazari A, Shekari F. et al. Hydrogel-mediated sustained systemic delivery of mesenchymal stem cell-derived extracellular vesicles improves hepatic regeneration in chronic liver failure. ACS Appl Mater Interfaces. 2019;11:37421-33
104. Haga H, Yan IK, Takahashi K, Matsuda A, Patel T. Extracellular vesicles from bone marrow-derived mesenchymal stem cells improve survival from lethal hepatic failure in mice. Stem Cells Transl Med. 2017;6:1262-72
105. Zhou Y, Liu S, Zhao M, Wang C, Li L, Yuan Y. et al. Injectable extracellular vesicle-released self-assembling peptide nanofiber hydrogel as an enhanced cell-free therapy for tissue regeneration. J Control Release. 2019;316:93-104
106. Eirin A, Zhu XY, Puranik AS, Tang H, McGurren KA, van Wijnen AJ. et al. Mesenchymal stem cell-derived extracellular vesicles attenuate kidney inflammation. Kidney Int. 2017;92:114-24
107. Zou X, Zhang G, Cheng Z, Yin D, Du T, Ju G. et al. Microvesicles derived from human Wharton's Jelly mesenchymal stromal cells ameliorate renal ischemia-reperfusion injury in rats by suppressing CX3CL1. Stem Cell Res Ther. 2014;5:40
108. Zou X, Gu D, Zhang G, Zhong L, Cheng Z, Liu G. et al. NK cell regulatory property is involved in the protective role of MSC-derived extracellular vesicles in renal ischemic reperfusion injury. Hum Gene Ther. 2016;27:926-35
109. Go V, Bowley BGE, Pessina MA, Zhang ZG, Chopp M, Finklestein SP. et al. Extracellular vesicles from mesenchymal stem cells reduce microglial-mediated neuroinflammation after cortical injury in aged Rhesus monkeys. Geroscience. 2020;42:1-17
110. Chen SY, Lin MC, Tsai JS, He PL, Luo WT, Herschman H. et al. EP4 antagonist-elicited extracellular vesicles from mesenchymal stem cells rescue cognition/learning deficiencies by restoring brain cellular functions. Stem Cells Transl Med. 2019;8:707-23
111. Riazifar M, Mohammadi MR, Pone EJ, Yeri A, Lässer C, Segaliny AI. et al. Stem cell-derived exosomes as nanotherapeutics for autoimmune and neurodegenerative disorders. ACS Nano. 2019;13:6670-88
112. Ruppert KA, Nguyen TT, Prabhakara KS, Toledano Furman NE, Srivastava AK, Harting MT. et al. Human mesenchymal stromal cell-derived extracellular vesicles modify microglial response and improve clinical outcomes in experimental spinal cord injury. Sci Rep. 2018;8:480
113. Drommelschmidt K, Serdar M, Bendix I, Herz J, Bertling F, Prager S. et al. Mesenchymal stem cell-derived extracellular vesicles ameliorate inflammation-induced preterm brain injury. Brain Behav Immun. 2017;60:220-32
114. Zhang S, Teo KYW, Chuah SJ, Lai RC, Lim SK, Toh WS. MSC exosomes alleviate temporomandibular joint osteoarthritis by attenuating inflammation and restoring matrix homeostasis. Biomaterials. 2019;200:35-47
115. Cosenza S, Toupet K, Maumus M, Luz-Crawford P, Blanc-Brude O, Jorgensen C. et al. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics. 2018;8:1399-410
116. Zhang S, Chuah SJ, Lai RC, Hui JHP, Lim SK, Toh WS. MSC exosomes mediate cartilage repair by enhancing proliferation, attenuating apoptosis and modulating immune reactivity. Biomaterials. 2018;156:16-27
117. Bier A, Berenstein P, Kronfeld N, Morgoulis D, Ziv-Av A, Goldstein H. et al. Placenta-derived mesenchymal stromal cells and their exosomes exert therapeutic effects in Duchenne muscular dystrophy. Biomaterials. 2018;174:67-78
118. Figliolini F, Ranghino A, Grange C, Cedrino M, Tapparo M, Cavallari C. et al. Extracellular vesicles from adipose stem cells prevent muscle damage and inflammation in a mouse model of hind limb ischemia: role of neuregulin-1. Arterioscler Thromb Vasc Biol. 2020;40:239-54
119. Lo Sicco C, Reverberi D, Balbi C, Ulivi V, Principi E, Pascucci L. et al. Mesenchymal stem cell-derived extracellular vesicles as mediators of anti-inflammatory effects: endorsement of macrophage polarization. Stem Cells Transl Med. 2017;6:1018-28
120. Zhao H, Shang Q, Pan Z, Bai Y, Li Z, Zhang H. et al. Exosomes from adipose-derived stem cells attenuate adipose inflammation and obesity through polarizing M2 macrophages and beiging in white adipose tissue. Diabetes. 2018;67:235-47
121. Liu H, Liang Z, Wang F, Zhou C, Zheng X, Hu T. et al. Exosomes from mesenchymal stromal cells reduce murine colonic inflammation via a macrophage-dependent mechanism. JCI Insight. 2019;4:e131273
122. Mathew B, Ravindran S, Liu X, Torres L, Chennakesavalu M, Huang CC. et al. Mesenchymal stem cell-derived extracellular vesicles and retinal ischemia-reperfusion. Biomaterials. 2019;197:146-60
123. Silva AKA, Perretta S, Perrod G, Pidial L, Lindner V, Carn F. et al. Thermoresponsive gel embedded with adipose stem-cell-derived extracellular vesicles promotes esophageal fistula healing in a thermo-actuated delivery strategy. ACS Nano. 2018;12:9800-14
124. Li X, Liu L, Yang J, Yu Y, Chai J, Wang L. et al. Exosome derived from human umbilical cord mesenchymal stem cell mediates miR-181c attenuating burn-induced excessive inflammation. EBioMedicine. 2016;8:72-82
125. Kordelas L, Rebmann V, Ludwig AK, Radtke S, Ruesing J, Doeppner TR. et al. MSC-derived exosomes: a novel tool to treat therapy-refractory graft-versus-host disease. Leukemia. 2014;28:970-3
126. Nassar W, El-Ansary M, Sabry D, Mostafa MA, Fayad T, Kotb E. et al. Umbilical cord mesenchymal stem cells derived extracellular vesicles can safely ameliorate the progression of chronic kidney diseases. Biomater Res. 2016;20:21
127. Xie H, Wang Z, Zhang L, Lei Q, Zhao A, Wang H. et al. Extracellular vesicle-functionalized decalcified bone matrix scaffolds with enhanced pro-angiogenic and pro-bone regeneration activities. Sci Rep. 2017;7:45622
128. Takafuji Y, Hori M, Mizuno T, Harada-Shiba M. Humoral factors secreted from adipose tissue-derived mesenchymal stem cells ameliorate atherosclerosis in Ldlr-/- mice. Cardiovasc Res. 2019;115:1041-51
129. Roccaro AM, Sacco A, Maiso P, Azab AK, Tai YT, Reagan M. et al. BM mesenchymal stromal cell-derived exosomes facilitate multiple myeloma progression. J Clin Invest. 2013;123:1542-55
130. Katakowski M, Buller B, Zheng X, Lu Y, Rogers T, Osobamiro O. et al. Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett. 2013;335:201-4
131. Tan JL, Lau SN, Leaw B, Nguyen HPT, Salamonsen LA, Saad MI. et al. Amnion epithelial cell-derived exosomes restrict lung injury and enhance endogenous lung repair. Stem Cells Transl Med. 2018;7:180-96
132. Zhou Y, Li P, Goodwin AJ, Cook JA, Halushka PV, Chang E. et al. Exosomes from endothelial progenitor cells improve the outcome of a murine model of sepsis. Mol Ther. 2018;26:1375-84
133. Zhou Y, Li P, Goodwin AJ, Cook JA, Halushka PV, Chang E. et al. Exosomes from endothelial progenitor cells improve outcomes of the lipopolysaccharide-induced acute lung injury. Crit Care. 2019;23:44
134. Chaput N, Théry C. Exosomes: immune properties and potential clinical implementations. Semin Immunopathol. 2011;33:419-40
135. Robbins PD, Morelli AE. Regulation of immune responses by extracellular vesicles. Nat Rev Immunol. 2014;14:195-208
136. Rhys HI, Dell'Accio F, Pitzalis C, Moore A, Norling LV, Perretti M. Neutrophil microvesicles from healthy control and rheumatoid arthritis patients prevent the inflammatory activation of macrophages. EBioMedicine. 2018;29:60-9
137. Ashrafian F, Shahriary A, Behrouzi A, Moradi HR, Keshavarz Azizi Raftar S, Lari A. et al. Akkermansia muciniphila-derived extracellular vesicles as a mucosal delivery vector for amelioration of obesity in Mice. Front Microbiol. 2019;10:2155
138. Zhang D, Lee H, Wang X, Rai A, Groot M, Jin Y. Exosome-mediated small RNA delivery: a novel therapeutic approach for inflammatory lung responses. Mol Ther. 2018;26:2119-30
139. Shang Y, Sun Y, Xu J, Ge X, Hu Z, Xiao J. et al. Exosomes from mmu_circ_0001359-modified ADSCs attenuate airway remodeling by enhancing FoxO1 signaling-mediated M2-like macrophage activation. Mol Ther Nucleic Acids. 2020;19:951-60
140. Bei Y, Das S, Rodosthenous RS, Holvoet P, Vanhaverbeke M, Monteiro MC. et al. Extracellular vesicles in cardiovascular theranostics. Theranostics. 2017;7:4168-82
141. Wei Z, Qiao S, Zhao J, Liu Y, Li Q, Wei Z. et al. miRNA-181a over-expression in mesenchymal stem cell-derived exosomes influenced inflammatory response after myocardial ischemia-reperfusion injury. Life Sci. 2019;232:116632
142. Gallet R, Dawkins J, Valle J, Simsolo E, de Couto G, Middleton R. et al. Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur Heart J. 2017;38:201-11
143. Han C, Zhou J, Liang C, Liu B, Pan X, Zhang Y. et al. Human umbilical cord mesenchymal stem cell derived exosomes encapsulated in functional peptide hydrogels promote cardiac repair. Biomater Sci. 2019;7:2920-33
144. Lee ST, Im W, Ban JJ, Lee M, Jung KH, Lee SK. et al. Exosome-based delivery of miR-124 in a Huntington's disease model. J Mov Disord. 2017;10:45-52
145. Haney MJ, Klyachko NL, Zhao Y, Gupta R, Plotnikova EG, He Z. et al. Exosomes as drug delivery vehicles for Parkinson's disease therapy. J Control Release. 2015;207:18-30
146. Haney MJ, Zhao Y, Harrison EB, Mahajan V, Ahmed S, He Z. et al. Specific transfection of inflamed brain by macrophages: a new therapeutic strategy for neurodegenerative diseases. PLoS One. 2013;8:e61852
147. Cooper JM, Wiklander PB, Nordin JZ, Al-Shawi R, Wood MJ, Vithlani M. et al. Systemic exosomal siRNA delivery reduced alpha-synuclein aggregates in brains of transgenic mice. Mov Disord. 2014;29:1476-85
148. Izco M, Blesa J, Schleef M, Schmeer M, Porcari R, Al-Shawi R. et al. Systemic exosomal delivery of shRNA minicircles prevents parkinsonian pathology. Mol Ther. 2019;27:2111-22
149. Kalani A, Chaturvedi P, Kamat PK, Maldonado C, Bauer P, Joshua IG. et al. Curcumin-loaded embryonic stem cell exosomes restored neurovascular unit following ischemia-reperfusion injury. Int J Biochem Cell Biol. 2016;79:360-9
150. Yang J, Zhang X, Chen X, Wang L, Yang G. Exosome mediated delivery of miR-124 promotes neurogenesis after ischemia. Mol Ther Nucleic Acids. 2017;7:278-87
151. Chen Z, Wang H, Xia Y, Yan F, Lu Y. Therapeutic potential of mesenchymal cell-derived miRNA-150-5p-expressing exosomes in rheumatoid arthritis mediated by the modulation of MMP14 and VEGF. J Immunol. 2018;201:2472-82
152. Yu L, Yang F, Jiang L, Chen Y, Wang K, Xu F. et al. Exosomes with membrane-associated TGF-β1 from gene-modified dendritic cells inhibit murine EAE independently of MHC restriction. Eur J Immunol. 2013;43:2461-72
153. Lv LL, Feng Y, Wu M, Wang B, Li ZL, Zhong X. et al. Exosomal miRNA-19b-3p of tubular epithelial cells promotes M1 macrophage activation in kidney injury. Cell Death Differ. 2020;27:210-26
154. Li ZL, Lv LL, Tang TT, Wang B, Feng Y, Zhou TL. et al. HIF-1α inducing exosomal microRNA-23a expression mediates the cross-talk between tubular epithelial cells and macrophages in tubulointerstitial inflammation. Kidney Int. 2019;95:388-404
155. Lv LL, Feng Y, Wen Y, Wu WJ, Ni HF, Li ZL. et al. Exosomal CCL2 from tubular epithelial cells is critical for albumin-induced tubulointerstitial inflammation. J Am Soc Nephrol. 2018;29:919-35
156. Wang B, Yao K, Huuskes BM, Shen HH, Zhuang J, Godson C. et al. Mesenchymal stem cells deliver exogenous microRNA-let7c via exosomes to attenuate renal fibrosis. Mol Ther. 2016;24:1290-301
157. Vandewalle J, Luypaert A, De Bosscher K, Libert C. Therapeutic mechanisms of glucocorticoids. Trends Endocrinol Metab. 2018;29:42-54
158. Wu H, Fan H, Shou Z, Xu M, Chen Q, Ai C. et al. Extracellular vesicles containing miR-146a attenuate experimental colitis by targeting TRAF6 and IRAK1. Int Immunopharmacol. 2019;68:204-12
159. Ran N, Gao X, Dong X, Li J, Lin C, Geng M. et al. Effects of exosome-mediated delivery of myostatin propeptide on functional recovery of mdx mice. Biomaterials. 2020;236:119826
160. Xie F, Zhou X, Fang M, Li H, Su P, Tu Y. et al. Extracellular vesicles in cancer immune microenvironment and cancer immunotherapy. Adv Sci (Weinh). 2019;6:1901779
161. Yan W, Jiang S. Immune cell-derived exosomes in the cancer-immunity cycle. Trends Cancer. 2020;6:506-17
162. Watson DC, Bayik D, Srivatsan A, Bergamaschi C, Valentin A, Niu G. et al. Efficient production and enhanced tumor delivery of engineered extracellular vesicles. Biomaterials. 2016;105:195-205
163. Choo YW, Kang M, Kim HY, Han J, Kang S, Lee JR. et al. M1 macrophage-derived nanovesicles potentiate the anticancer efficacy of immune checkpoint inhibitors. ACS Nano. 2018;12:8977-93
164. Go G, Lee J, Choi DS, Kim SS, Gho YS. Extracellular vesicle-mimetic ghost nanovesicles for delivering anti-inflammatory drugs to mitigate gram-negative bacterial outer membrane vesicle-induced systemic inflammatory response syndrome. Adv Healthc Mater. 2019;8:e1801082
165. Wan Z, Zhao L, Lu F, Gao X, Dong Y, Zhao Y. et al. Mononuclear phagocyte system blockade improves therapeutic exosome delivery to the myocardium. Theranostics. 2020;10:218-30
166. Gangadaran P, Hong CM, Ahn BC. An update on in vivo imaging of extracellular vesicles as drug delivery vehicles. Front Pharmacol. 2018;9:169
Author contact
![]() Corresponding author: Bi-Cheng Liu (E-mail: liubc64com).
Corresponding author: Bi-Cheng Liu (E-mail: liubc64com).